
chronique du diamat n°3
"Intelligent design" ou force autoconservatrice
Prenons l’un des arguments actuels du créationnisme au sérieux… tant il pointe ce qu’on pourrait qualifier de défaillance gradualiste du darwinisme. La théorie très américaine de l’Intelligent design critique sévèrement ce trait évolutionniste sur la base d’observations d’ordre biochimique principalement. Constatant par exemple que toute enzyme possède une complexité « irréductible » dans sa structure et par suite dans son fonctionnement, sa spécificité catalytique, les antidarwiniens affirment qu’on ne saurait expliquer par un processus graduel d’évolution « par petites touches » (mutations génétiques ponctuelles sur « le gène » de l’enzyme) l’extrême minutie de la mécanique intégrée d’une enzyme quelle qu’elle soit. L’exemple fameux est celui du flagelle bactérien, petite merveille de rotor qui la rend mobile en milieu aqueux, et pourtant réputé primitif par rapport à ceux des eucoryotes : on peut en voir un aperçu dans la vidéo ci-dessous.

Les antidarwiniens affirment qu’on ne peut faire émerger « par étapes », accidentelles de surcroît, aucune machinerie même la plus élémentaire. Une tapette à souris par exemple ne peut se concevoir que finalisée, jamais comme l’aboutissement d’un processus darwinien graduel de sélection naturelle. Si cette tapette à souris est composée d’une planchette, d’un fil de fer armé d’un pic et d’un ressort, il est impossible que la planchette ait émergée seule d’abord pour ensuite se munir accidentellement d’un ressort lui-même inutile avant l’apparition accidentelle du fil de fer armé finalisant le mécanisme.
A cela les darwiniens gradualistes répliqueront qu’une molécule peut évoluer progressivement dans son fonctionnement au fil des changements de circonstances, par accumulation et superposition de fonctions opportunes se complétant ou se remplaçant. Mais ce sont là les limites du gradualisme, qui à ce jour ne trouve pas de modèle pertinent à opposer aux partisans du créationnisme, si ce n’est à modifier en amont leur théorie de la connaissance par une sorte de pseudo-humilité empirique, toujours plus relativiste : « la science n’est pas faite pour dire le vrai » mais elle est ce qui est communément admis à l’instant t, ce qui est « réfutable », etc.
Cette faiblesse vient très clairement de la part du camp darwinien d’un manque de logique dialectique, autrement dit d’un excès de logique formelle, de « mécanisme ». L’évolution du vivant ne procède pas par « un » mécanisme, mais par l’accumulation de nombreux mécanismes superposés avec le temps dirigés pour accomplir une tendance : celle de lutter contre les changements, aussi paradoxal que cela paraisse. Il existe au niveau des cellules vivantes plusieurs propriétés permettant de diminuer drastiquement la fréquence des mutations, parce que celles-ci sont quasi-systématiquement néfastes au bon fonctionnement de l’ensemble. Or le perfectionnement progressif de ces mécanismes a abouti à des métabolismes qui, pour tenter de diminuer l’impact de changements environnementaux sur l’espèce, peuvent « anticiper » ces changements avec des contre-tendances. Autrement dit « changer pour ne rien changer », ce qui est impossible, la preuve étant que la vie n’a finalement jamais cessé d’évoluer ! On peut dire que l’évolution dialectique du vivant est une série de changements profonds fondés sur des mécanismes sélectionnés pour s’opposer à tout changement... La première pierre de cet édifice étant la propriété autoréplicative des ARN et ADN à l'origine de l’histoire de la vie, c'est-à-dire « avant » la sélection naturelle d’une certaine façon.
Cette approche reste darwinienne dans le sens où tous ces mécanismes ont été eux mêmes conservés et accumulés par la sélection naturelle. Mais elle relativise le modèle darwinien dans le sens où l’évolution est une conséquence de propriétés fondamentales du vivant (lutter contre les changements par des changements à contre-courant), et non une simple « caractéristique » du vivant parmi d’autres (se nourrir, se reproduire, etc.), qui n’a fait que développer et perfectionner ces modalités moléculaires primitives de « conservation » (contre-mouvement permanent plutôt que fixité).
Un exemple de cette tendance auto-conservatrice du vivant est celui de la Rubisco, l’enzyme la plus abondante dans toute la biosphère, présente chez tous les végétaux pour incorporer le carbone atmosphérique et créer leur matière organique (matière carbonée), assurant leur croissance et celle consécutive des herbivores, des carnivores, etc.
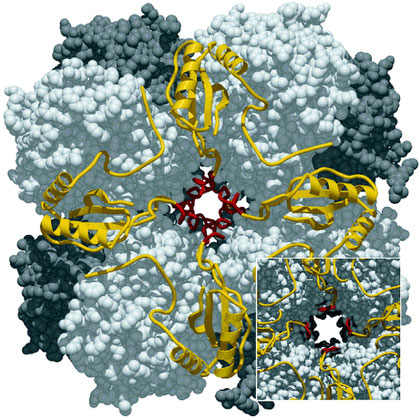
La Ribulose biphosphate carboxylase
Quelques préalables avant de poursuivre.
La photosynthèse est apparue très tôt dans l’histoire de la vie, faisant émerger des formes capables de s’autoproduire, c'est-à-dire de créer de la matière organique à partir de matière minérale (eau, dioxyde de carbone, sels minéraux). Cette faculté fut sans doute centrée sur l’apparition de cette enzyme fondamentale, la Ribulose biphosphate carboxylase ou Rubisco : enzyme capable d’ajouter un CO2 atmosphérique à un sucre à 5 carbones (le ribulose 1,5 biphosphate) pour préparer la formation de sucres à 6 carbones (glucose, fructose, etc.) à travers un processus de recyclage du carbone appelé le cycle de Calvin, au niveau des chloroplastes des cellules végétales chlorophylliennes.
Or cette carboxylation, qui est une « réduction » chimique du CO2 atmosphérique en matière organique carbonée, ne peut être que couplée à une « oxydation » donneuse d'électrons; celle de l’eau transformée en dioxygène sous l’effet de l’énergie lumineuse via les pigments chlorophylliens. Autrement dit, dans l’atmosphère primitive terrestre qui contenait beaucoup de CO2 et qui ne contenait pas d’O2 (personne ne « respirait » encore à l’époque), l’apparition des premiers organismes photosynthétiques ont créé et massivement rejeté comme un déchet inutile l’O2, tout en consommant une partie du CO2 atmosphérique… C’est là qu’on découvre un effet tout à fait intéressant sur le caractère profondément « contradictoire » et dialectique des grandes étapes évolutives qui ont dû suivre dans cette histoire du vivant.
En effet la rubisco a été sélectionnée pour son affinité au CO2, or à l’époque de sa sélection l’atmosphère primitive était encore, avons-nous dit, privée de O2. Quelques millions d’années plus tard les êtres photosynthétiques s’étaient tellement développés, avant l’apparition des premières formes capables de respirer (consommation d’O2), que l’atmosphère elle-même s’en était enrichi.
Il se trouve que la rubisco a aussi une certaine affinité pour l’O2, loin d’être négligeable : un sucre en C5 sur cinq subit une oxygénation plutôt qu’une carboxylation par la rubisco, ce qui est une perte sèche pour la plante. Ce handicap n’a donc pas pu s’exprimer au moment de la « fixation » précoce de la rubisco dans les premiers génomes chlorophylliens. Mais alors pourquoi, depuis, cette enzyme n’a-t-elle pas évolué, comme le prévoit le modèle gradualiste darwinien, vers une perte d’affinité pour l’O2, une meilleure affinité pour le CO2?
Curieusement la Rubisco n’a quasiment pas changé au cours du temps, gage de son efficacité stratégique dans les mécanismes hautement complexes de la photosynthèse. A ce degré d’importance métabolique, tout s’opposait au moindre changement et le handicap, manifesté trop tardivement, fut maintenu, malgré son énorme « coût » en matière et en énergie…
Les organismes chlorophylliens n’en sont toutefois pas resté là puisque des stratégies de contournement ont pu être acquises par la suite pour tenter de limiter les effets du handicap de la rubisco. Ainsi la « photorespiration » a été pour les plantes une manière de limiter les dégâts occasionnés par la perte sèche en carbone (20%), en recyclant le sucre en C5 oxygéné (alcool toxique s’il s’accumule) pour recharger au moins trois de ces carbones dans le cycle de Calvin créateur de matière organique, et alimenter opportunément avec les autres carbones des anabolismes secondaires (la création d’acides aminés en particulier). Cette photorespiration fonctionne comme un « moindre mal » mais ne remet pas en cause l’affinité malheureuse de la rubisco pour l’O2. C’est donc une façon de "botter en touche" face à la contradiction dialectique opposant la carboxylation liée à la rubisco au dioxygène libéré qu’elle provoque mais qui finit par la géner (« lutte et unité des contraires » selon Lénine).
D’autres réponses ont été plus impressionnantes, qui n’ont pourtant pas non plus annulé le handicap fondamental de la rubisco. De nombreuses plantes dites en « C4 » ou en « CAM » ont innové en faisant apparaître en plus de la rubisco une autre enzyme de carboxylation, la PEP carboxylase, sans affinité pour l’O2, et qui peut « stocker » massivement le CO2 pour le libérer autour des rubisco en amont de leur activité de façon à désavantager l’O2 environnant. En effet le rendement de la rubisco dépend du rapport environnant entre [CO2] et [O2]. Plus le milieu est concentré en CO2 par rapport au taux fixe du O2 dissout (comme dans l'atmosphère primitive), plus il carboxylera et moins il oxygénera. La libération d’un stock de CO2 massif autour des rubisco est donc pour ces plantes une façon d’abaisser au maximum la coûteuse photorespiration (qui baisse le bilan en CO2 fixé et augmente la dépense énergétique). Ce mécanisme a d’ailleurs d’autres avantages, dont celui de favoriser la fixation du carbone en climat chaud et sec (puisque le CO2 est moins soluble donc moins présent autour des rubisco quand il fait chaud).
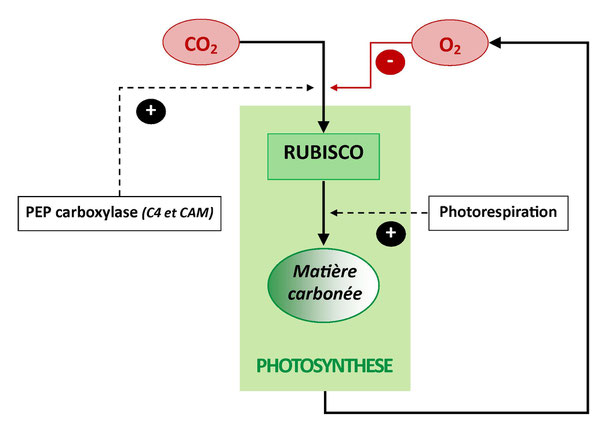
Une contradiction dialectique entre carboxylation et oxygénation au sein de la RUBISCO
Il est donc très interessant de voir à travers cet exemple d’enzyme fondamentale de la photosynthèse, quoiqu’handicapée, la capacité du vivant de développer des stratégies limitant ce handicap, de façon tout à fait darwinienne, tout en conservant l’enzyme elle-même ! La force conservatrice si puissante qu’elle a ici « fossilisé » notre enzyme telle qu’elle fonctionnait dans l’atmosphère primitive sans O2, a vaincu la force qu’on appelle pression de sélection et qui est pourtant d’après les néodarwiniens la force centrale de l’évolution des espèces. Deux forces s’opposent, l’une (la conservatrice) domptant quelquefois l’autre (la sélection naturelle) ou s’y accommodant quand cela est opportun. Voilà qui donne à la "stabilité" apparente du vivant une toute autre dimension que celle donnée en première instance par les créationnistes...