
Chronique du diamat n°1
Sur les conflits entre symbiotes
Dialectique de la Nature compte sans doute parmi les plus grands classiques du marxisme, bien qu’Engels son auteur, l’eût considéré comme une ébauche inachevée et dont la réalisation « demandera[it] encore beaucoup de temps » (Lettre à Marx, 30 mai 1873). Plus d’un siècle de lutte idéologique plus tard, naturalistes, généticiens, physiologistes résistent encore : Il serait absurde, voire dogmatique de considérer que « la science de la nature en est arrivée au point de ne plus pouvoir échapper à la synthèse dialectique », comme l’affirmait Engels dans l’Anti-Düring. C’est l’heure de l’éclipse des « menaces » bolcheviks sur le « monde libre », l’heure du pragmatisme libéral, de la méfiance vis-à-vis des théories, enfin libéré du « totalitarisme » marxiste qui régna au vingtième siècle jusque dans les amphithéâtres des universités françaises, pourtant il ne suffirait pas d’un seul Engels pour mener cette colossale synthèse dialectique tant les dernières découvertes sont nombreuses et la capacité de les intégrer théoriquement étroite.
C’est que le champs de la biologie, du point de vue du dialecticien, apparaît assez contradictoire : c’est bien sur celui de l’évolution des espèces, fondation du matérialisme sur laquelle Marx et Engels ont installé, littéralement, l’histoire de l’Homme lui-même, celle mouvante de la lutte des classes. Mais c’est aussi le régne d’une apparente fixité, qui invite tout naturaliste à la description fascinée autant que stérile. Le complexe d’infériorité de la jeune biologie face à la physique ou à la chimie, chez qui l’inexorable mouvement dialectique, entropique, est à ce jour largement admis, vient sans doute de cette mauvaise habitude, depuis les origines, de considérer la Nature comme un cabinet de curiosité dans lequel la vie même serait par définition un objet inatteignable, profondément mystérieux. En réalité même la révolution darwinienne ne permit pas de réaliser jusqu’au bout cette prise de conscience des lois fondamentales de la dialectique en biologie, en reconnaissant que la Vie, comme toute autre manifestation de la matière, répond à une "contradiction [c'est à dire une] présence dans un phénomène d'aspects opposés s'excluant mutuellement mais qui, en même temps se supposent l'un l'autre et sont inséparables dans un phénomène donnée" comme le disait Kark Marx.
Or c’est précisément « en portant au devant du caractère dialectique de ces lois [naturelles] la conscience des lois de la pensée dialectique » (Dialectique de la Nature) qu’on mesure leur véritable potentiel pour comprendre le vivant dans son mouvement permanent, précisément parcequ’il y est paradoxalement contre-intuitif : la dialectique du vivant est en effet une dialectique entre mouvement et… stabilité. Une dialectique de l’équilibre instable plutôt que du mouvement pur et mécanique.
Dans son infinie complexité de voies évolutives, convergeant toutes dans la tendance (toujours finalement inversée en son contraire) à se « conserver » malgré les changements environnementaux, autrement dit à se transformer pour susbister, le vivant est une des manifestations de la matière qui dépasse et nie son entropie même par un contre-mouvement « stabilisateur », auto-correcteur, incapable d’équilibre durable. Une telle tendance engendre elle-même des formes atypiques particulièrement étudiées des comtemplateurs naturalistes, sur lesquelles le dialecticien se penchera avec tout autant de profit ; celle des « impasses » évolutives qu’on appelle coévolution. La coévolution est en effet la forme la plus aboutie, et en même temps paradoxalement la plus instable, d’un cas d’école de la contradiction dialectique s’approfondissant jusqu’au non-sens. Elle illustre parfaitement la célèbre loi de la « lutte ET de l’unité des contraires » (la première et la plus connue des trois lois de la dialectique) tout en démontrant l’absence de finalité dans la direction, pourtant tangible (celle d’une tendance à l’autoconservation sans cesse anihilée) de l’évolution du vivant.
Si la vie cherche pour se conserver plus efficacement (lutte pour la survie) à échapper, à s’émanciper du milieu, par définition instable, plutôt que de s’y adapter étroitement, au moyen de stratégies métaboliques toujours plus complexes, le développement du piège de « la lutte et de l’unité des contraires » atteint son paroxysme quand une espèce devient le milieu d’une autre et réciproquement. Ainsi des figuiers du genre Ficus et de leurs hymenoptères pollinisateurs Agaonidae.
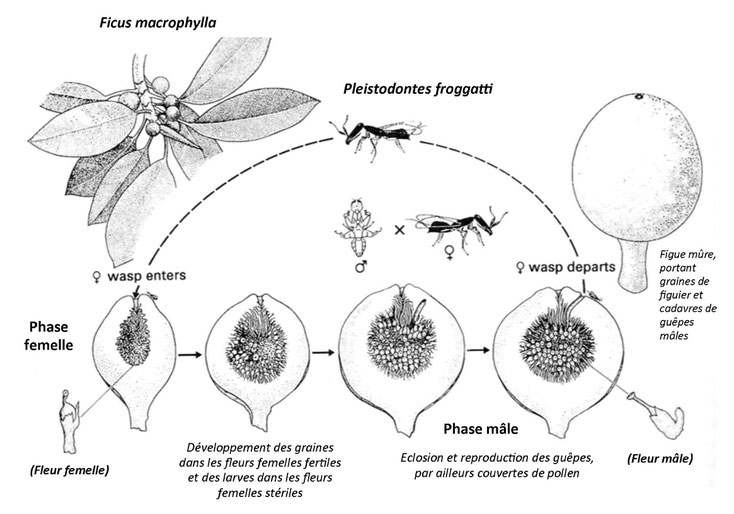
Figure 1: Le cycle de vie d'un Ficus monoïque (chaque plante est bisexuée) et de son pollinisateur
Il existe une stricte dépendance entre plus de sept cents espèces de Ficus et autant d’espèces de guêpes pollinisatrices. Cette interdépendance se traduit par une coadaptation très poussée, dont les éléments décrivent ce qu’on appelle une contradiction dialectique antagoniste, avec un pôle unité particulièrement puissant, puisque deux forces auto-conservatrices sont dirigées ici l’une contre l’autre, plutôt que chacune contre leur milieu changeant. Dans chaque couple de cette famille, le figuier a un pollinisateur exclusif sans lequel il ne peut plus se reproduire, tandis que le pollinisateur ne peut se reproduire lui-même que dans les figues qu’il pollinise.
La figue est une inflorescence en forme d’urne fermée par un ostiole préformé ne laissant passer qu’une sorte de guèpe, et contient potentiellement trois types de minuscules fleurs (mâles, femelles fertiles et femelles stériles). Déjà fécondée par un mâle, la guèpe entrée dans une figue en est désormais définitivement prisonnière: elle se met à pondre dans les fleurs femelles stériles. Celles-ci sont adaptées à recevoir les œufs qui s’y développent en larves des deux sexes (style court de même taille que l’oviscape des guèpes contrairement aux longs styles des fleurs fertiles) et ne donneront jamais de graines. Les guèpes écloses se reproduiront entre elles au sein de la figue toujours fermée. Les mâles aptères mourront sur place, non sans avoir aidé les femelles, à la fois fécondées et portant le pollen des fleurs mâles désormais mûres, à sortir de la figue mûre (plus sèche et friable avec le temps) pour aller pondre dans une autre, plus jeune et non encore inséminée, sur un autre figuier.
Les Ficus dioïques, c'est-à-dire dont les plantes sont soit mâles soit femelles, présentent une complication intéressante du système : Une figue de plante femelle contiendra uniquement des fleurs femelles fertiles, et devra être inseminée par des guèpes incapables d’y pondre leurs œufs (absence de fleurs femelles stériles à style court). Une figue de plante mâle contiendra au contraire un mélange de fleurs mâles couvrant de pollen les guèpes écloses d’une insemination de fleurs femelles stériles à style court coexistant avec les fleurs mâles. Ces figues mâles ne porteront donc pas de graines fécondées, mais des larves de guèpes uniquement. Entre le Ficus dioïque et la guèpe, on peut donc identifier l’unité consistant en une interdépendance reproductive absolue, et en même temps une opposition, un conflit puisqu’une figue mâle sera frustrée (pas de graines) en faisant naître des guèpes, tandis qu’une figue femelle frustrera la guèpe chargée de pollen qui y sera piègée (pas de ponte possible).
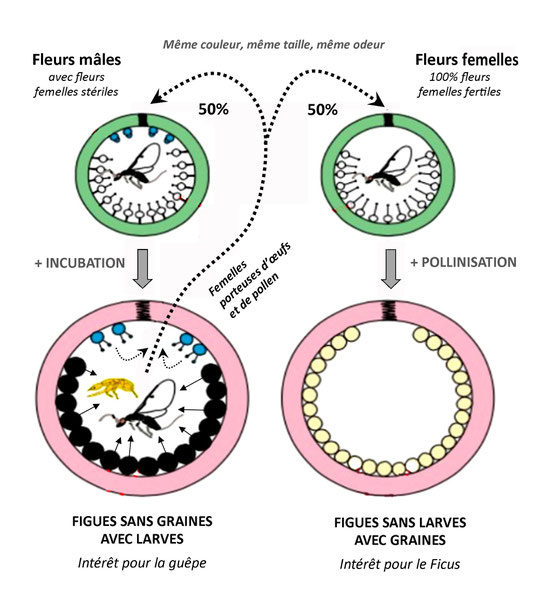
Figure 2: Une contradiction dialectique entre le Ficus dioïque et son pollinisateur
Chaque protagoniste a développé des adaptations anatomiques pourtant coûteuses et dirigées dans le sens de la satisfaction du partenaire : c’est l’aspect unité. Par exemple, les pollinisateurs présentent un abdomen segmenté qui se dégonfle à l’air libre et sec, protégeant le pollen sous les bords superposés des segments rétractés y compris à l’entrée traumatique dans un ostiole de figue à féconder. Certains ont même développé des poches à pollen spécialisées au niveau de l’abdomen qu’ils remplissent activement avant d’entrer dans une nouvelle figue. De son coté le figuier « sacrifie » une partie de ses fleurs femelles (stériles) pour la ponte des guèpes, en raccourssissant leur style à la taille des oviscapes des guèpes mères (chez les figuiers mâles donc).
En revanche, les figuiers résolvent le conflit d’intérêt relatif avec les guèpes fécondées pollinisatrices en brouillant les pistes : toutes les figues, qu’elles soient mâles (ponte possible pour la guèpe mais pollinisation inutile) ou femelles (pollinisation mais ponte impossible), ont strictement la même couleur et le même marqueur olfactif pour les guèpes, de sorte qu’une guèpe fécondée et chargée de pollen soit incapable de « choisir » la figue qui satisferait immédiatement ses intérêts (la figue mâle à fleurs femelles stériles).
Ce genre de contradiction dialectique est assez typique et pourrait fournir un bon exemple illustrant la première loi hégélienne reformulée par Engels. Mais elle est d’abord chargée d’une signification particulière à la biologie, car il n’existe à cette coévolution aucune échappatoire salutaire sous forme de saut qualitatif (comme dans le cas d’une contradiction dialectique antagoniste) : les guèpes de la même famille devenues « parasites » du Ficus et capable avec un oviscape démesuré, de pondre dans plusieurs figues de l’extérieur et sans y entrer, sont elles-mêmes dépendantes du couple coadapté, car en l’absence de l’espèce pollinisatrice prisonnière du jeu, le parasite disparaîtrait lui aussi faute de reproduction des figuiers à terme.

Figure 3: Une guêpe parasite dotée d'un long oviscapte de ponte, échappant à l'emprisonnement de la figue.
En réalité, ce type de coadaptation extrême est sans doute la forme la moins stable, la moins efficace des tentatives du vivant pour se réadapter en permanence au milieu changeant, car il multiplie les inféodations et ferait disparaître immédiatement deux espèces à la moindre inadaptation. Il attise la curiosité mais s’avère sans doute beaucoup moins « évolué » que les espèces dites « généralistes », celles qui se satisfont indifféremment de nombreux milieux, de nombreuses conditions changeantes, sans devoir véritablement changer en permanence, comme le font les couples en coévolution, dans cette course effrénée à reculons pour se répondre à l’infini sans aucune autre finalité que celle d’avoir le dernier mot.
Oui, en ce sens on peut dire que l’évolution du vivant n’a pas de finalité, comme le martèlent les néodarwiniens relativistes adaptes du buissonement, puisque ce type de coévolution, marginal en fait, montre qu’on peut entrer dans l’impasse d’un dialogue « absurde », sans fin et vulnérable, tout autant que dans une issue évolutive relativement émancipatrice vis-à-vis d’une contrainte environnementale (cas général). Mais il faut sortir dialectiquement la loi de la marginalité de ses exceptions et reconnaître aussi que de telles impasses sans finalité soulignent par l’absurde, en négatif, l’autofinalité du vivant dans ses grands traits évolutifs, ceux qui ont permis une certaine émancipation du milieu, des poissons aux mammifères placentaires par exemple, une certaine capacité d’anticipation des changements de milieu avec les découvertes récentes en épigénétique, modalités d’évolution rapide en période de bouleversement environnemental, etc. Autant de raisons pour les Engels de demain de renvoyer dos à dos idéalistes finalistes et antifinalistes, et d’élaborer enfin une vision cohérente du phénomène vivant incluant son évolution comme une loi interne nécessaire plutot qu’une vague propriété parmi d’autres.